



Swanandi Gawde, M. Optom
Assistant Professor, Lotus College of Optometry, Mumbai, India
In the human eyeball, photoreceptor cells, rods and cones, are responsible for colour vision. Rod cells are for night vision, and cone cells are for colour vision. The visible spectrum of the human eyeball ranges from 380 to 750 nm.
Photoreceptors and Colour Processing
Colour vision exploration starts with photoreceptors that primarily perceive the colours red, green, and blue. The normal colour vision possesses three retina cone cells termed L, M, and S. These cones contain different photo-pigments sensitive to various wavelengths. Signals from these cones are combined into two retino‐geniculate mechanisms: the so‐called red-green and blue-yellow cone‐opponent channels. Their absorption peaks correspond to 560, 530, and 425 nm, respectively. (1)
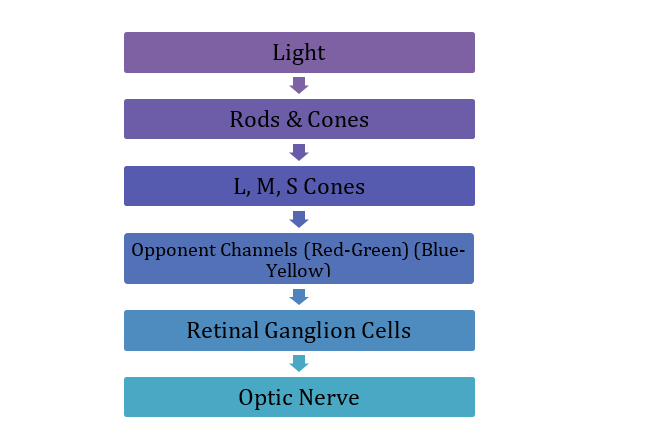

Figure 1: The flowchart highlighting the colour processing pathway.
The retinal ganglion cells transmit colour information to the brain. And photoreceptor cells adjust their sensitivity according to how much they are stimulated. (2) As the amount of long-wavelength light in the overall illumination increases, the red receptors will reduce their sensitivity and so maintain a stable output. (3)
| Rod Cells | Cone Cells |
|---|---|
| Night Vision | Colour Vision |
| Highly sensitive to light | Less sensitive |
| No colour detection | Detect colours |
| Mainly the peripheral retina | Concentrated in fovea |
| Active in dim light | Active in daylight |
Table 1: This table highlights the difference between the rod and cone cells.
Converging evidence from studies using psychophysical methods and visual evoked potentials suggests that infants red, green colour mechanism develop first, and the blue-yellow mechanism develops around 4 to 8 weeks later, with infants being trichromatic (both cone-opponent mechanisms are active) by 3 months. (4)
How the Brain Maintains Colour Stability
The Retinex Theory (Retina + Cortex) proposed by Edwin Land, suggests that the brain calculates surface colour by independently comparing the lightness of an object across three different, separate channels (one for each cone type). The cortex evaluates the scene and effectively “subtracts” the illumination to determine the reflectance of the surface. (5) Colour constancy is the tendency of human eyes to perceive colour of a surface as remaining constant despite changes in the spectrum of the illumination. (6) Colour constancy is a key feature of the Visual neuron 4 region. It contains functions related to object recognition. For example, an apple remains red both in light and in shadow. Colour and brightness constancy is also found in V1 and V2, and the effects are especially prominent in V4. (7,8)
The cells adjust their colour-tuning function in response to changes in the chromaticity of the illuminant. This adjustment corresponds with psychophysical perception. (9)
| Object | Morning Light | Evening Light | Perceived Colour |
|---|---|---|---|
| Apple | Warm light | Cool shadow | Red |
| Traffic Signal | Sunlight | Rainy weather | Red |
| Safety Sign | Daylight | Artificial light | Same colour |
| Banana | Kitchen light | Outdoor light | Yellow |
Table 2: This Table shows the real-world examples of colour constancy.
Conclusion
Colour constancy is a remarkable example of perceptual constancy, where the visual system transforms variable sensory input into stable perception. Like size constancy, which allows objects to appear the same size despite changes in viewing distance, colour constancy enables consistent recognition of objects in changing environments. Although colour is ultimately a perceptual construct, the sophisticated processing mechanisms of the brain ensure that our visual world remains coherent, reliable, and meaningful.
References
- Nathans, J. (1999). The evolution and physiology of human colour vision: insights from molecular genetic studies of visual pigments. Neuron, 24(2), 299–312.
- Zhang, B., Zhang, R., Zhao, J., Yang, J., & Xu, S. (2024). The mechanism of human colour vision and potential implanted devices for artificial colour vision. Frontiers in Neuroscience, 18, 1408087.
- Smithson, H. E. (2005). Sensory, computational and cognitive components of human colour constancy. Philosophical Transactions of the Royal Society B: Biological Sciences, 360(1458), 1329–1346.
- Teller, D. Y. (1998). Spatial and temporal aspects of infant colour vision. Vision Research, 38(21), 3275–3282. 10.1016/S0042-6989(97)00468-9.
- Land, E. H. (1977). The retinex theory of colour vision. Scientific American, 237(6), 108–129.
- Foster, D. H. (2011). Colour constancy. Vision Research, 51(7), 674–700.
- Conway, B. R. (2001). Spatial structure of cone inputs to colour cells in alert macaque primary visual cortex (V1). Journal of Neuroscience, 21, 2768–2783. 10.1523/JNEUROSCI.21-08-02768.2001.
- Zeki, S. (1983). The distribution of wavelength and orientation selective cells in different areas of the monkey visual cortex. Proceedings of the Royal Society B: Biological Sciences, 217, 449–470. 10.1098/rspb.1983.0020.
- Schein, S., & Desimone, R. (1990). Spectral properties of V4 neurons in the macaque. Journal of Neuroscience, 10, 3369–3389.
About the Author


Swanandi Gawde
Assistant Professor,